|
新闻动态
|
科研进展
昆明植物所在揭示杜鹃花属物种形成过程中基因流异质性与基因组结构关系方面取得重要进展 理解基因组结构如何塑造基因流在基因组的分布景观,是揭示植物物种形成机制的重要内容。近缘物种间的基因流既可能促进遗传多样性,也可能导致基因组的同质化。基因组结构包括基因组序列的整体排布及其特征,如基因密度、染色体倒位、转座子分布、着丝粒演化,以及染色体的数目和长度等,都被证实会以多种方式影响基因流在基因组上的分布。然而,染色体在三维空间中的结构与基因流之间是否存在关联,以及它们的相互关系如何进一步影响物种形成仍有待进一步探究。 杜鹃花属(Rhododendron L.)是北半球物种最丰富的木本属,全球有 1000 多种。喜马拉雅-横断山及邻近地区是杜鹃花属的现代分布和分化中心,包含 320 余种,其中 210 多种为特有种。自中新世以来,受造山运动和亚洲季风影响,该地区的杜鹃花属物种经历了快速辐射演化。近缘物种间生殖隔离较弱,自然杂交普遍,多样的生境与频繁杂交共同促进了物种分化。然而,杜鹃花属在频繁基因流作用下如何维持物种界限的遗传机制尚不清晰。 近日,中国科学院昆明植物研究所高连明(现山东农业大学)团队联合国内外团队,在 New Phytologist 发表了题为 The genomic architecture of introgression during Rhododendron speciation 的研究成果。研究以分布于中国西南山地及周边地区的杜鹃花属糙叶杜鹃亚组(R. subsect. Scabrifolia)8 种杜鹃花为研究对象,开展杜鹃花属物种形成过程中基因渐渗的基因组景观特征研究,发现染色体三维空间结构中的 A 区室与 B 区室在基因组分化程度上存在显著差异。其中,与着丝粒紧密相关的 B 区室通常表现出较高的遗传分化,而富含基因的 A 区室分化程度较低,并呈现更极端的 𝑓dm 值,表明局部基因渐渗更为广泛。研究提出,不同基因组区室之间基因流的异质性可能推动杜鹃花属的物种形成,并在植物基因流分布景观的塑造中发挥关键但尚未完全揭示的作用。 该研究首先利用二代和三代测序技术(NGS,HiC,HIFI)从头组装了爆杖花(R. spinuliferum)高质量的基因组(图 1),BUSCO 评估值为97.1%,全基因LAI 平均值为19.68。爆杖花基因组大小为634.5M,重复序列在基因组中占70.21%,基因组共预测到35,406 个蛋白编码基因。通过HiC数据的染色体三维结构分析,识别了染色体 A/B 区室,通常认为A 区室对应着常染色质,B 区室对应着异染色质。结果显示,染色体 A/B 区室的分布与基因密度、GC 含量及Gypsy 元件丰度等基因组结构特征高度吻合。随后,对横断山及其近邻的云贵高原地区分布的糙叶杜鹃亚组的8个物种15个居群共139个个体进行全基因组重测序,通过系统发育、群体遗传学和比较基因组学分析全面解析该多样性群体的基因组演化。
图1 爆杖花的基因组结构特征 糙叶杜鹃亚组的演化背景与基因流历史 基于系统发育分析结果,虽然在不同数据集之间存在核基因树冲突和核质冲突,但整个亚组可以明确地分为两个分支,即SSS 分支(包括糙叶杜鹃R. scabrifolium,碎米花R. spiciferum 和爆杖花)和HMPR 分支(包括粉背碎米花R. hemitrichotum,柔毛碎米花R. mollicomum,柔毛杜鹃R. pubescens 和腋花杜鹃R. racemosum)。基于叶绿体基因组估算的分化时间表明,糙叶杜鹃亚组在大约3.85百万年前开始分化。流式细胞术测定的基因组大小表明,核质系统发育强烈冲突的两个物种(富源杜鹃R. fuyuanense 和腋花杜鹃)均为四倍体。基于SNP 的群体结构分析显示,8 种杜鹃花均形成了独立的遗传谱系。多种基因流检测(包括D 值,f4-ratio,f-branch 及QuIBL)结果均支持物种间存在频繁的基因交流。种群历史模拟的结果显示物种形成期间的基因流多为间歇性、不连续的。综上所述,糙叶杜鹃亚组的8个物种分布区部分重叠,分化时间较短,分化过程中存在基因流,但均形成了独立的遗传谱系。
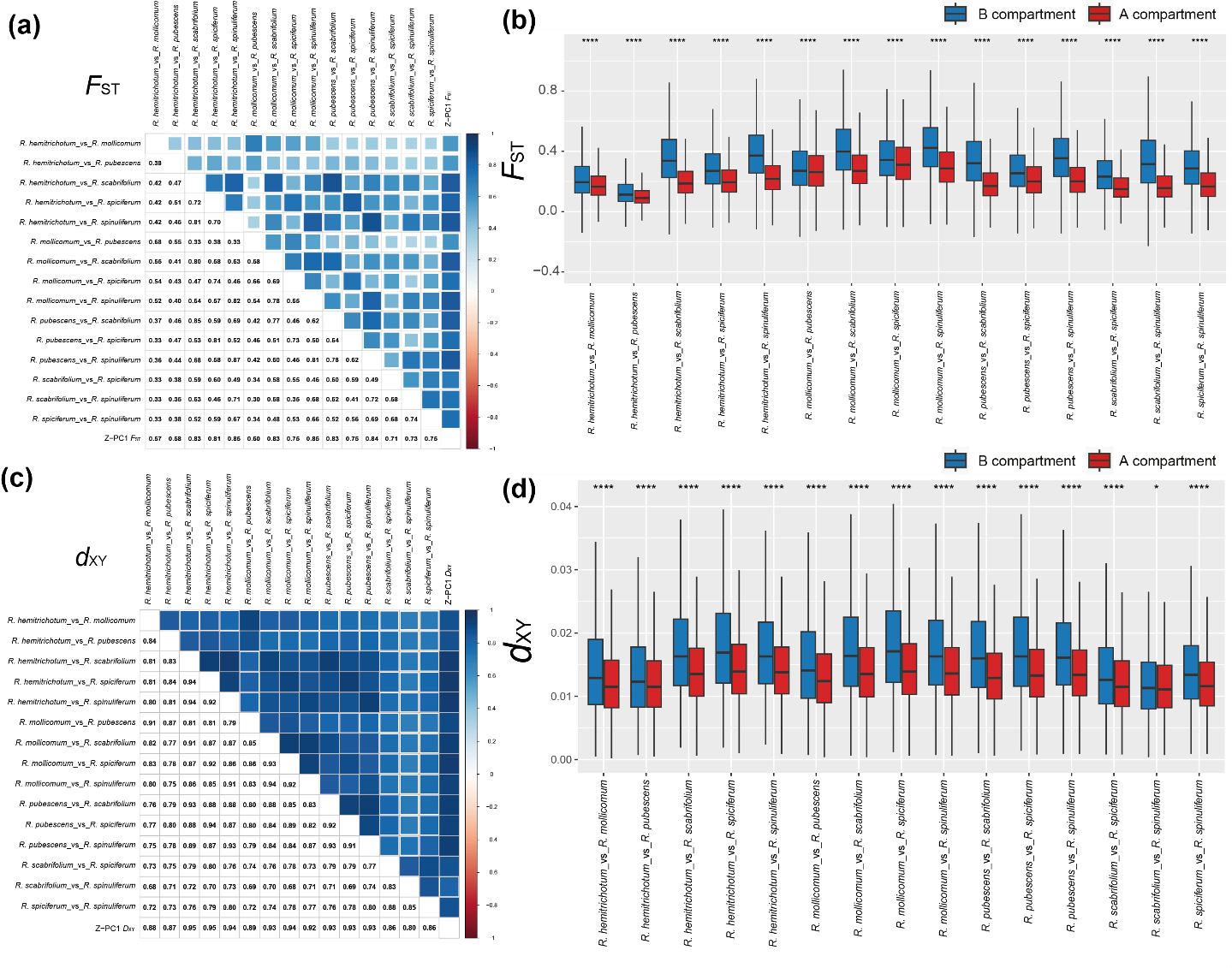
图2 糙叶杜鹃亚组八个物种的分布、群体结构及演化历史 基因组结构是遗传多样性与分化差异的基础 该研究发现,在糙叶杜鹃亚组物种的自然群体中,染色体 A 区室与 B 区室在基因组分化和基因流强度上存在显著差异(图 3)。结果显示包含着丝粒的B 区室在物种间表现出更大的遗传分化(𝐹𝑆𝑇)和分异(𝐷𝑋𝑌 ),相比之下,A 区室则在不同物种中倾向于低的遗传分化水平,具有较低的遗传分化和分异。基于沿基因组10 kb 滑窗的基因流分析表明,在几乎所有的𝑓dm 组合中,A 区室前5% 的𝑓dm 平均值比B 区室前5% 的𝑓dm 平均值高,说明在A 区室中具有更大程度的局部基因渐渗(图 4)。基因组异质性景观的形成可能源于基因组结构对重组率、突变率和选择压力的综合影响:B 区室靠近着丝粒,着丝粒进化迅速且易发生倒位,低重组率加上转座元件积累和杂合不相容性等都会抑制基因流;而 A 区室基因丰富、重组率高,使有益等位基因更易渗入并通过适应性选择快速固定。总体来看,基因组结构不仅塑造了染色体不同区室的遗传多样性与遗传分化模式,也在物种形成和适应性进化中发挥重要作用。
图3 10 kb的基因组滑窗下,A/B 区室间物种的遗传分化值(𝐹𝑆𝑇)及遗传分异值(𝐷𝑋𝑌)的差异
图4 10 kb的基因组滑窗下,A/B 区室间物种的基因流分布差异 生态因素与基因组结构共同影响物种辐射演化 该研究表明,糙叶杜鹃亚组的物种演化受到生态因素与基因组结构的共同作用。在横断山地区,晚中新世的造山活动和气候变化为物种形成提供了微生态与演化机遇。更新世气候波动和冰期—间冰期循环可能促使原本隔离的物种扩展分布,从而在迁移期间发生间歇性基因流。在空间隔离期,整个基因组,尤其是 B 区室发生分化,积累新突变并筛选祖先单倍型,促进物种沿着不同的演化方向发展;而在发生基因流时期,A 区室易于基因渐渗,使近缘物种间形成遗传上的动态网络,快速响应环境变化。研究推测,不同染色体区室可能倾向于保存不同来源的适应性变异:B 区室的新突变通过长期固定积累,影响基因组稳定性并引导演化方向;而 A 区室的共享等位基因或适应性基因渐渗则促进物种快速适应环境。总体来看,生态因素与基因组结构的交互作用共同塑造了糙叶杜鹃亚组物种的快速适应性演化。 综上所述,在糙叶杜鹃亚组物种间的全基因组范围内,遗传分化值(𝐹𝑆𝑇)和遗传分异值(𝐷𝑋𝑌)呈现显著差异。这种差异在染色体的 A 区室与 B 区室间表现出一致性:B 区室在物种间通常分化程度较高,而 A 区室分化程度较低,可能因物种间频繁的基因流所致。研究推测,间歇性的基因流结合着丝粒的快速进化,是驱动杜鹃花属物种辐射演化的内部动力。B 区室紧密关联着丝粒,为物种分化提供了多样的进化方向;而 A 区室的遗传多样性与环境异质性则有助于物种形成以及对环境变化的快速适应。 中国科学院昆明植物研究所已毕业博士研究生秦汉韬(现任西南林业大学讲师)为论文第一作者,高连明教授(现山东农业大学)、李洪涛研究员和李德铢教授(现山东农业大学)为论文共同通讯作者。英国爱丁堡大学Alex D. Twyford 博士和 Richard I. Milne 博士、埃克塞特大学 Hans-Wilhelm Nützmann 博士、西双版纳热带植物园严丽君副研究员,昆明植物所已出站的博士后郑伟博士,以及在读博士研究生莫智琼、朱明姝等参与了此项研究。该研究得到国家自然科学基金、国家重点研发计划和云南省基础研究专项重大项目的资助。 |

版权所有 Copyright © 2002-2025 中国科学院昆明植物研究所,All Rights Reserved 【滇ICP备05000394号】
地址:中国云南省昆明市蓝黑路132号 邮政编码:650201
点击这里联系我们